Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional
DOI: https://dx.doi.org/10.19137/huellas-2025-2913
Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional
ARTÍCULOS
El uso de tecnologías de la información geográfica en el estudio de la elección de árboles dormideros por monos aulladores negros y dorados (Alouatta caraya)
The use of geographic information technologies in the study of sleeping tree selection by black and gold howler monkeys (Alouatta caraya)
O uso de tecnologias de informação geográfica no estudo da escolha de árvores-dormitório por bugios-pretos e dourados (Alouatta caraya)
Félix Ignacio Contreras[1]
Universidad Nacional del Nordeste / Consejo Nacional de Investigaciones Científicas y Técnicas
ignaciocontreras@exa.unne.edu.ar
Rocío Débora Giles[2]
Universidad Nacional del Nordeste / Consejo Nacional de Investigaciones Científicas y Técnicas
Martín Kowalewski[3]
Universidad Nacional del Nordeste / Consejo Nacional de Investigaciones Científicas y Técnicas
RECIBIDO 13-05-2025 / ACEPTADO 01-08-2025
Cita sugerida: Contreras, F. Giles, R. D. y Kowalewski, M. (2025). El uso de tecnologías de la información geográfica en el estudio de la elección de árboles dormideros por monos aulladores negros y dorados (Alouatta caraya), Revista Huellas, Volumen 29, Nº 2, Instituto de Geografía, EdUNLPam: Santa Rosa. Recuperado a partir de: http://cerac.unlpam.edu.ar/index.php/huellas
Resumen
La comprensión de distribución y patrones espaciales constituyen uno de los principales principios de la Geografía, tanto en variables humanas como en naturales. En este contexto, aplicar herramientas de Tecnologías de Información Geográfica permiten potenciar, como en este caso, estudios de comportamiento de la especie de monos carayá (Alouatta caraya) a la hora de seleccionar árboles dormideros a lo largo del año. Con este trabajo, se pudo comprobar que una tropa de monos ubicados en un Bosque frente al Parque Provincial San Cayetano (Corrientes), opta por ocupar árboles de mayor porte y/o sectores con mayor volumen. Teniendo en cuenta los sitios dormideros registrados y las condiciones meteorológicas para aquellos días, se pudo concluir que la selección de estos lugares se encuentra asociado a la búsqueda de un confort climático, más que una necesidad por la búsqueda de alimentos o protección de depredadores.
Palabras clave: Distribución; Biogeografía; TIG; Corrientes; Alouatta caraya
Abstract
Understanding distribution and spatial
patterns is one of the core principles of Geography, applicable to both human and natural variables. Within
this context, the application of Geographic Information Technologies provides a valuable tool for enhancing
studies of species behaviors such as, in this case, the sleeping tree selection by black howler monkeys
(Alouatta caraya) throughout the year.
This study confirmed that a troop of monkeys located in a forest adjacent to San Cayetano Provincial Park
(Corrientes, Argentina) tends to occupy larger trees and/or areas with greater vegetative volume.
Considering the recorded sleeping sites and the corresponding meteorological conditions, it was concluded
that the selection of these locations is primarily associated with the pursuit of climatic comfort, rather
than driven by proximity to food sources or protection from predators.
Keywords: Distribution; Biogeography; GIS; Corrientes; Alouatta caraya
Resumo
A compreensão da
distribuição e dos padrões espaciais constitui um dos princípios fundamentais da
Geografia, tanto para variáveis humanas quanto naturais. Nesse contexto, a aplicação de
ferramentas de Tecnologias de Informação Geográfica permite potencializar estudos de
comportamento de espécies, como no presente caso, referente à seleção de
árvores-dormitório pela espécie de bugios-pretos (Alouatta caraya) ao longo do ano. Este estudo
comprovou que um grupo de macacos localizado em uma floresta em frente ao Parque Provincial San Cayetano
(Corrientes, Argentina) tende a ocupar árvores de maior porte e/ou setores com maior volume vegetal.
Considerando os locais de pernoite registrados e as condições meteorológicas
correspondentes, concluiu-se que a escolha desses espaços está principalmente associada
à busca por conforto térmico, mais do que à proximidade de alimentos ou
proteção contra predadores.
Palavras-chave: Distribuição; Biogeografia; TIG; Corrientes; Alouatta caraya
Introducción[4]
La selección de sitios y árboles dormideros es un comportamiento clave en la ecología de los primates, la cual puede estar influenciada por una variedad de factores sociales, ecológicos, fisiológicos, antrópicos y de salud que inciden directamente en su supervivencia y bienestar (Anderson 1998; Day et al. 1999; Di Bitetti et al. 2000; Brotcorne et al. 2014; Brividoro et al. 2019, Li et al. 2022). En particular, los monos aulladores negros y dorados (Alouatta caraya), una de las especies más distribuidas del género Alouatta en Sudamérica, exhiben patrones complejos en la elección de sus dormideros, los cuales podrían responder a múltiples presiones adaptativas (Kowalewski & Zunino 2005; Brividoro et al. 2019).
El mono aullador es la especie del género Alouatta con la distribución más amplia y meridional (Di Fiore et al., 2010; Oklander et al., 2019). Se encuentra en Paraguay, el sur de Brasil, el este de Bolivia y el norte de Argentina, llegando hasta la provincia de Santa Fe, así como en el norte de Uruguay (Zunino, 2004; Di Fiore y Campbell, 2007; Jardim et al., 2020). Es una especie de primate no humano caracterizada por una dieta folívora-frugívora y comportamientos estrictamente arbóreos que se encuentran en grupos sociales cohesivos de hasta 20 individuos entre adultos, juveniles e infantes (Oklander et al., 2019, Kowalewski et al., 2019).
Durante el descanso nocturno, los monos carayá utilizan los árboles altos, con copas grandes y ramas que ofrecen un sustrato seguro para dormir en áreas asociadas a las últimas sesiones de alimentación diarias durante el anochecer y en secuencias para evitar repeticiones posiblemente para evitar contacto con parásitos (Zunino 1986; Kowalewski & Zunino 2005). Brividoro et al., (2019) sugiere para la misma especie que el patrón de usos de aboles dormideros ofrecen una buena estabilidad, evitación de parásitos, acceso a fuentes de alimento y defensa de áreas de acción intergrupal). Por otro lado, la termorregulación nocturna representa un desafío particular para especies arbóreas que duermen expuestas a las condiciones ambientales, y podría influir significativamente en la selección de árboles que brinden mayor protección térmica (Eppley et al., 2017; Fei et al., 2019). Este factor es importante para evaluar en las decisiones de selección de árboles dormideros por aulladores, especialmente en ambientes con marcada estacionalidad térmica como las áreas de distribución para la especie en el noreste de Argentina. Así, los primates pueden utilizar los árboles dormideros para termorregulación, seleccionando sitios que les brinden protección contra condiciones climáticas extremas y promuevan la conservación de energía (Ellison et al., 2019; Mekonnen et al., 2021; Mills et al., 2021). Esto implica elegir árboles que les brinden refugio de la lluvia, el viento y el sol, y también permitan comportamientos de agrupamiento cohesivo para reducir la pérdida de calor.
En este contexto, el uso de tecnologías de la información geográfica se presenta como una herramienta para estudiar el comportamiento de los primates de forma no invasiva y eficiente (Gazagne et al., 2025). Así, contar con información georreferenciada acerca de la ubicación de los árboles dormideros en complemento con información, como la altura de los árboles obtenida mediante el uso de drones, permite no solo inferir de manera remota los posibles motivos de su elección, sino predecir la composición mínima necesaria de un bosque para sostener grupos de aulladores negros y dorados. Ante lo expuesto, este trabajo tiene como objetivo aplicar herramientas de tecnologías de información geográfica, para explorar factores asociados al confort térmico en la selección de árboles por Alouatta caraya desde una perspectiva integradora. Así nuestras a fin de determinar si existe algún patrón en la selección de árboles dormideros por monos aulladores negros y dorados relacionados a las variaciones de temperaturas estacionales que les permita obtener beneficios termoregulatorios.
Área de estudio
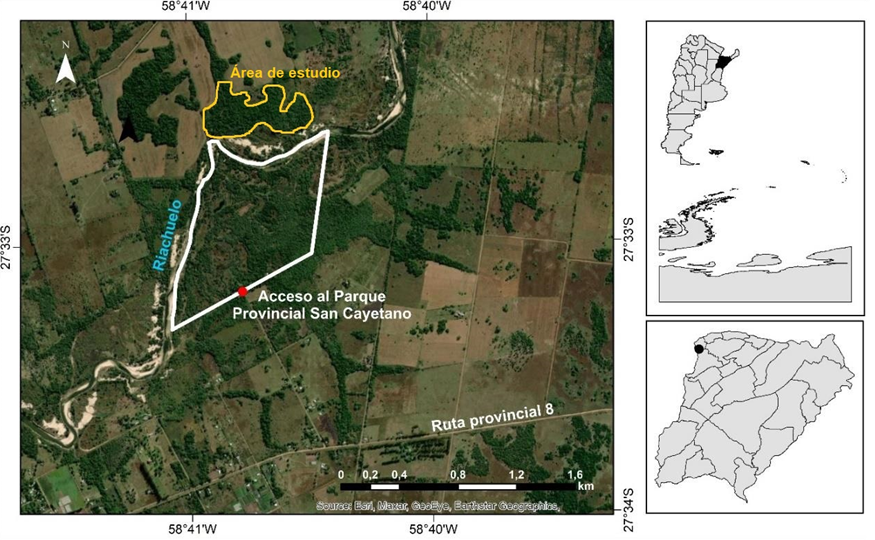
El estudio se realizó en un bosque en galería semideciduo ubicado frente al Parque Provincial San Cayetano y la Estación Biológica de Corrientes (27°30’ S, 58°41’ O), dentro de la cuenca del arroyo Riachuelo, en el noroeste de la provincia de Corrientes, Argentina. Cabe destacar que esta formación boscosa forma parte de los 24 fragmentos de bosque que albergaban grupos de monos aulladores negros y dorados (Alouatta caraya). Además, el paisaje corresponde a una terraza no inundable del arroyo Riachuelo (Contreras et al., 2022), donde coexisten bosques altos y bajos, sabanas con palmeras, pastizales y vegetación acuática vinculada a diversos cuerpos de agua (Figura N° 1).
Obtención del modelo digital de elevaciones (MDE) y el ortomosaico de fotografías aéreas
El MDE y el ortomosaico fueron generados mediante un vuelo del DJI Mini 4 Pro que cuenta con un sensor CMOS de 1/1,3 pulgadas, píxeles efectivos: 48 MP. Por último, se realizó el procesamiento de las fotografías aéreas con el software Global Mapper 25.1. La generación de un MDE de alta precisión se logró luego de la concreción de una secuencia de etapas o fases que a continuación se detallan.
Primera Fase: Planificación del Vuelo. Antes de ir a terreno se definieron los siguientes parámetros:
● Altura de vuelo: 100 m
● Solape frontal: 80 %
● Solape lateral: 50%
● Resolución espacial de las imágenes a obtener: 3,4 cm/px
● Velocidad promedio de vuelo: 30,0 km/h
● Fecha del vuelo: 17 de marzo de 2025. Para la elección del día se tuvo en cuenta la ausencia de nubosidad y evitar altas temperaturas que pudieran perjudicar al drone.
Segunda Fase: Corresponde al vuelo y captura de fotografías aéreas, cuya duración fue de 19 minutos y para lo que se utilizaron dos baterías.
Tercera Fase: Corresponde al posproceso que incluye desde la descarga de las imágenes obtenidas durante el vuelo hasta la generación del MDE. Luego de la importación de las imágenes en el software Global Mapper 25.1, se realizó la generación del MDE mediante el método de interpolación basado en la ponderación de distancia inversa que supone que los puntos más cercanos están más relacionados que otros mediante su función.
Cuarta Fase: La generación de cartografías se realizó mediante herramientas incluidas en Sistemas de información Geográfica, ArcGIS 10.5.
Toma de datos
Para el registro de datos comportamentales, se seleccionó un grupo de Alouatta carayaponer composición del grupo. La identificación individual de los monos se realizó considerando diferencias en el tamaño corporal y el dimorfismo sexual (Rumiz 1990; Kowalewski 2007), así como también mediante el uso de caravanas previamente colocadas en estudios anteriores y la presencia de cicatrices naturales. ¿El monitoreo de los sitios de descanso son sitios de descanso diario o solo dormideros nocturnos? Dejar siempre lo mismo sin cambian las preguntas se llevó a cabo utilizando un dispositivo GPS. Se registró la ubicación del grupo al amanecer, en el punto donde se encontraba durmiendo. Este procedimiento se repitió durante tres días consecutivos cada mes, a lo largo de un período de 12 meses.
Densidad de distribución dormideros
En ArcGIS 10.5, la densidad de Kernel, permite calcular la densidad de elementos, en las proximidades de estos, tanto para puntos como para líneas (Díaz et al., 2022), de la cual se han generado tres categorías: Alta, Media y Baja. Posteriormente se superpusieron los datos de densidad al MDE de modo de poder visualizar/identificar, la opción un determinar un rango de alturas en la elección de árboles dormideros.
Obtención de datos de temperatura del bosque
Los datos de las temperaturas y humedad relativa del bosque fueron registrados con un data logger Hobo Pro V2 U23-002, colocado a 1,5 m de altura en el centro de este, en dirección norte. El instrumental fue configurado para registrar un dato por hora entre el 15/09/2024 y el 05/03/2025.
Resultados y discusión
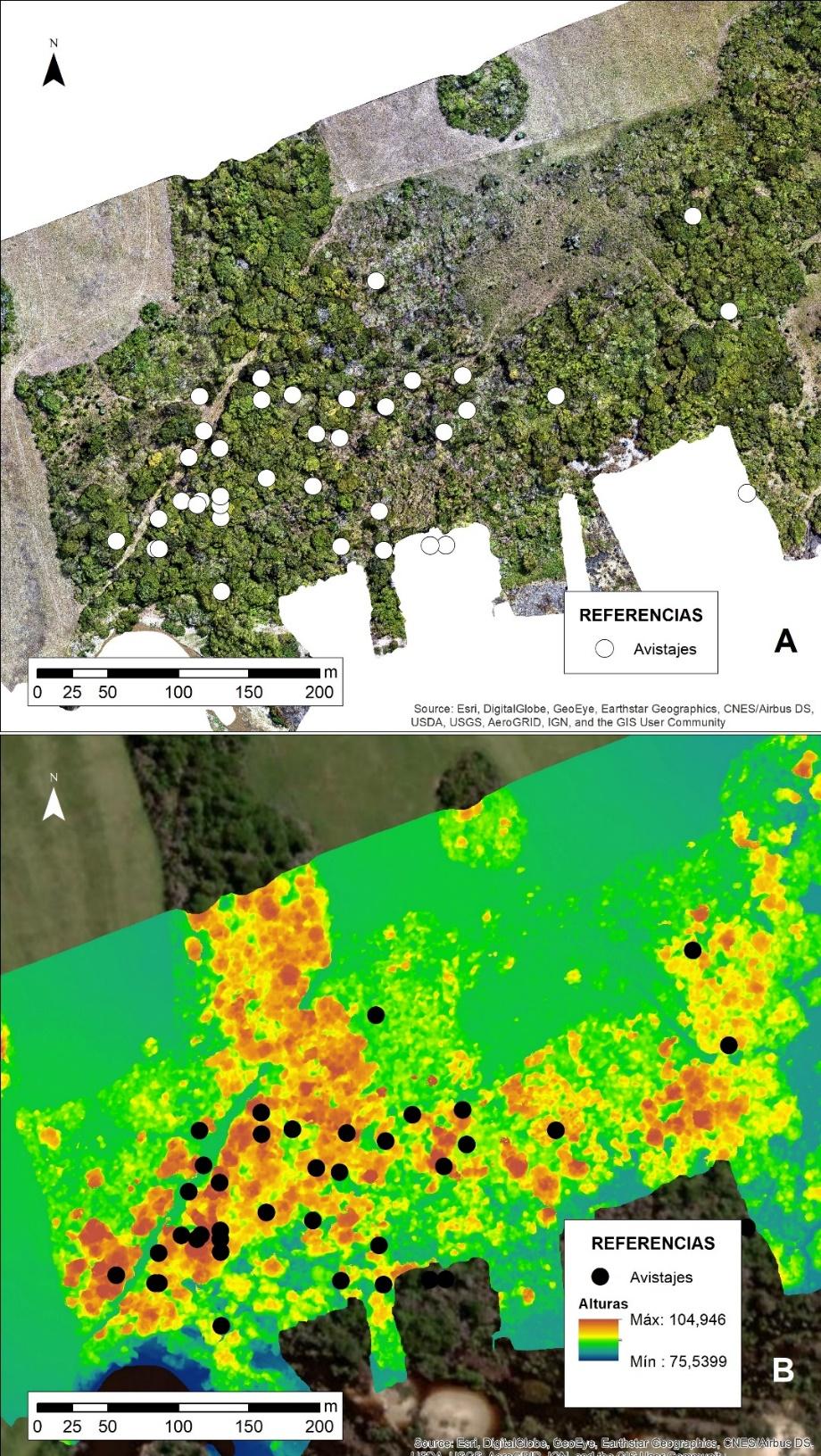
La generación del ortomosaico permite observar con gran resolución la distribución de los sitios dormideros del grupo de Alouatta caraya (Figura N° 2 A). En principio se observa la elección por la porción occidental del mismo, el cual concentra el 72 % de los casos, siguiendo la porción central con 21 % y el restante 8 % al oriental. Cabe destacar que la porción central presenta espacios más abiertos conformados, principalmente, por tacuarales (Guadua angustifolia).
Por otro lado, y si bien el área boscosa es una unidad alargada, a nivel de paisaje se encuentra segmentado de otros ambientes ya que, al sur se encuentra el arroyo Riachuelo y el resto se encuentra rodeado por pastizales. Por lo observado, estos últimos poseen una fuerte presión humana, tanto en el trazado de caminos como en la reducción de la biomasa, fácilmente comparable con la situación del pastizal central próximo al tacuaral.
Figura N° 2. A. Ortomosaico del bosque fecha 17 de marzo de 2025 y la distribución de los sitios dormideros. B. Modelo Digital de Elevaciones y la distribución de sitios dormideros
Fuente: Elaboración propia.
En cuanto a las alturas, y observando la Figura N° 2 B, se podría inferir que los monos optan por dormir en aquellos árboles de mayor altura, sin importar la sección del bosque. En este sentido, las alturas promedio de los árboles rondan entre los 25 y 30 m respecto del pastizal que los circunda.
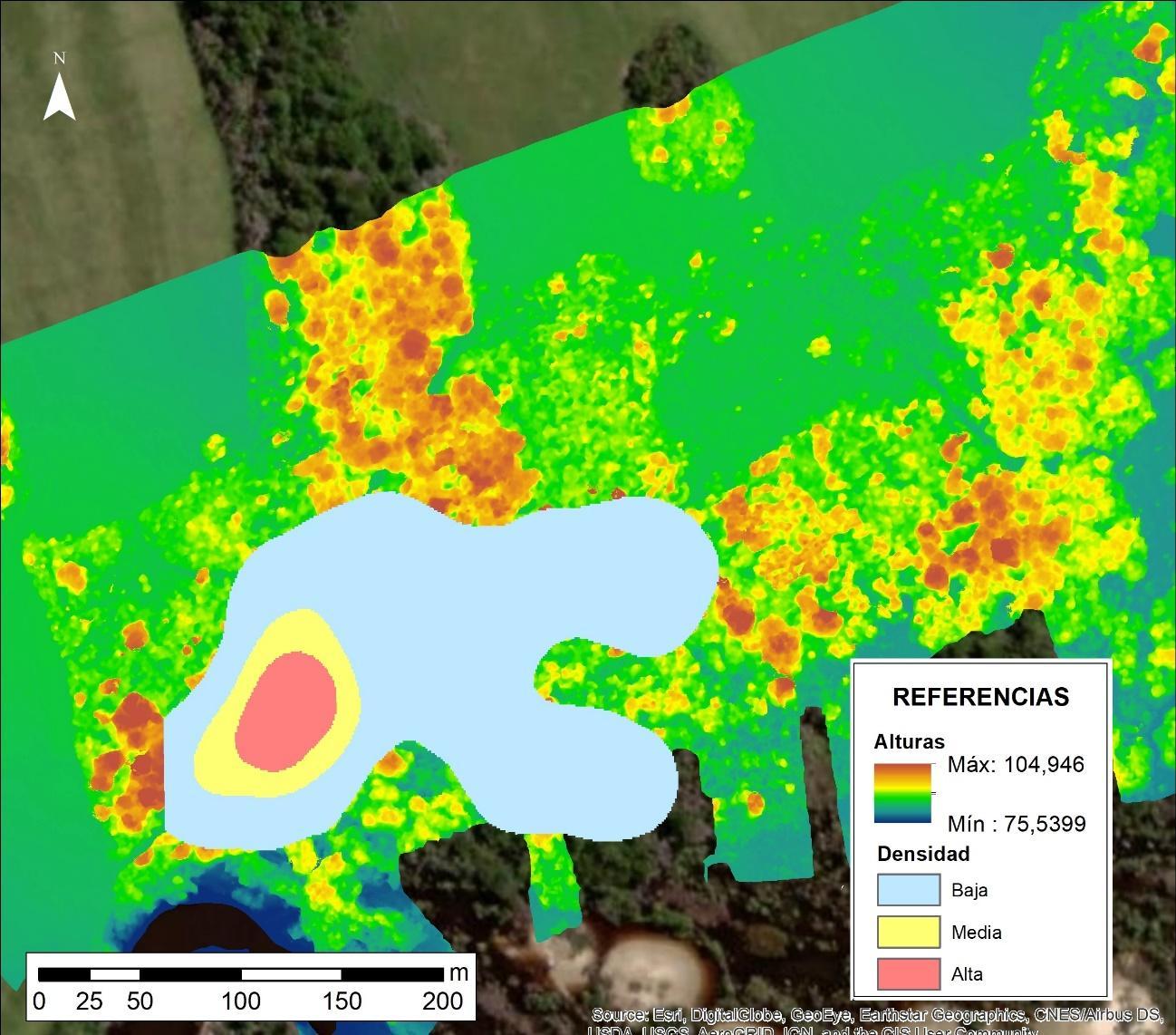
El análisis de densidad de Kernell de la Figura N° 3 se condice con lo descrito anteriormente, donde la mayor densidad de sitios dormideros se encuentra en la porción centro occidental, lugar donde existen las mayores alturas y mayor volumen boscoso. A partir de allí se desprenden dos corredores hacia el este, siendo el ubicado al norte el que conecta el occidente con el oriente.
Figura N° 3. Densidad de Kernel de la distribución de sitios dormideros de monos carayá en un bosque de la ribera del arroyo Riachuelo
Fuente Elaboración propia.
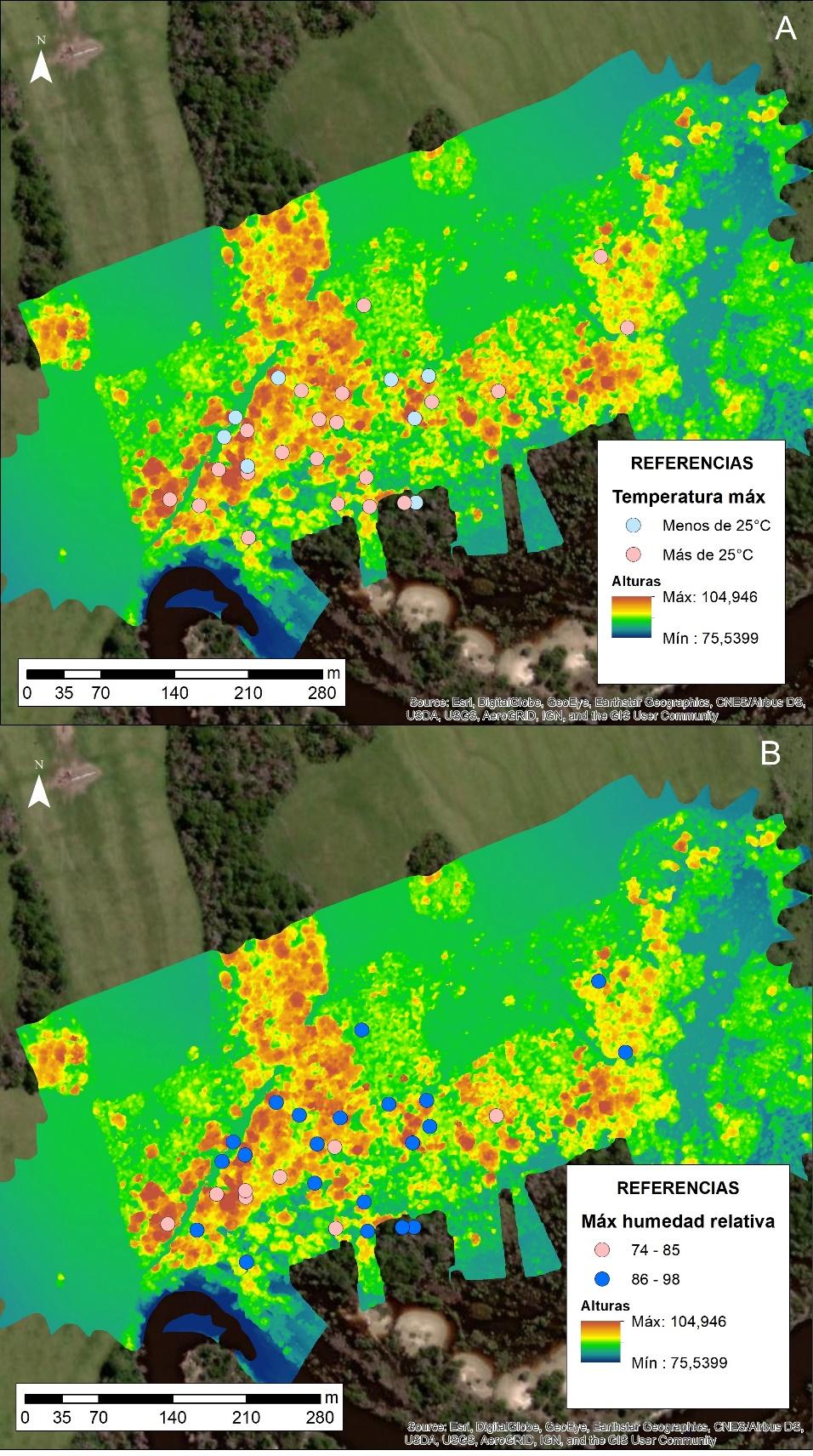
Respecto a la selección de sitios dormideros y las condiciones meteorológicas, en la Figura N° 4.A se observa que, cuando la temperatura máxima era superior a 25° C, hay una mayor distribución en árboles más bajos y sectores del bosque más abiertos (en promedio 2 m menos). Precisamente hay un mayor número de registros en las áreas menos densas. Por el contrario, cuando la temperatura máxima es inferior a 25° C, hay una mayor concentración en el sector más denso de puntos, el cual coincide con el mayor volumen de masa boscosa.
Este comportamiento, sería una respuesta en búsqueda de un confort térmico, ya que, en días de calor, los espacios más abiertos y de borde permiten el ingreso de ráfagas de viento y con ello disminuyendo la sensación de calor. Por el contrario, cuando las temperaturas son reducidas, la opción por áreas más densas se debe al microclima que ofrece este lugar, donde la humedad relativa es mayor y, por lo tanto, las temperaturas mínimas son menos extremas.
Con la humedad relativa habría una relación similar, es decir, cuanto mayor es la humedad relativa y sobre todo en días calorosos, la primera opción será buscar espacios más abiertos (Figura 4.B). La elección por estos lugares se puede dar en temperaturas inferiores a 25°C con humedades relativas superiores al 85 %. En noches frías, en cambio, los monos carayá tienden a seleccionar sitios de mayor volumen arbóreo, como árboles altos dentro del bosque, que ofrecen un microclima más cálido y estable. Aun así, en ciertos casos, podrían optar por ubicaciones elevadas que maximicen la exposición temprana a la radiación solar al amanecer, favoreciendo una recuperación térmica más rápida.
Figura N° 4. Sitios de elección de árboles dormideros. A. Valores de temperatura máxima durante el día. B. Valores de humedad relativa máxima durante el día. Bosque de ribera frente al Parque Provincial San Cayetano (Corrientes)
Fuente Elaboración propia.
En cuanto al viento y la lluvia, se ha observado que, con la llegada de un frente frío y precipitaciones, los monos, por lo general, optan por dormideros en árboles más bajos. Cabe destacar que las precipitaciones asociadas al viento sur se relacionan con tormentas del tipo frontal, las cuales pueden desarrollar fuertes ráfagas, granizo e importantes montos pluviométricos (Contreras, et al., 2020).
Cuadro N° 1. Datos de temperatura y humedad relativa dentro del bosque de ribera frente al Parque Provincial San Cayetano y dirección del viento y registro de precipitaciones. Período 22/12/2022-13/12/2023
|
Fecha |
Altura |
Temp Máx |
Temp Mín |
Temp Prom |
HR Máx |
HM Mín |
HR prom |
DV y RLl |
|
22/12/2022 |
90 |
28 |
19 |
36 |
81 |
26 |
49 |
|
|
6/1/2023 |
93 |
35 |
18 |
26 |
81 |
22 |
51 |
N |
|
7/1/2023 |
87 |
36 |
20 |
28 |
81 |
28 |
53 |
N |
|
8/1/2023 |
101 |
36 |
18 |
27 |
79 |
51 |
23 |
S |
|
9/1/2023 |
94 |
33 |
15 |
25 |
87 |
30 |
53 |
N |
|
6/2/2023 |
98 |
34 |
18 |
27 |
75 |
29 |
49 |
SE |
|
7/2/2023 |
101 |
38 |
21 |
30 |
75 |
26 |
49 |
N |
|
8/2/2023 |
<Null> |
39 |
24 |
31 |
74 |
28 |
52 |
N |
|
20/3/2023 |
89 |
31 |
22 |
26 |
98 |
61 |
84 |
N |
|
21/3/2023 |
92 |
32 |
23 |
27 |
94 |
51 |
80 |
N |
|
22/3/2023 |
93 |
32 |
23 |
27 |
95 |
56 |
83 |
N |
|
23/3/2023 |
85 |
32 |
23 |
27 |
95 |
55 |
80 |
S |
|
28/3/2023 |
<Null> |
27 |
16 |
22 |
93 |
52 |
77 |
S |
|
29/3/2023 |
100 |
29 |
17 |
24 |
93 |
52 |
78 |
N |
|
30/3/2023 |
88 |
32 |
22 |
28 |
97 |
63 |
81 |
N – LL |
|
31/3/2023 |
100 |
25 |
20 |
22 |
98 |
90 |
97 |
S |
|
2/6/2023 |
96 |
24 |
15 |
20 |
98 |
71 |
87 |
N |
|
3/6/2023 |
92 |
25 |
18 |
22 |
96 |
74 |
87 |
N |
|
4/6/2023 |
103 |
25 |
20 |
22 |
95 |
78 |
90 |
N |
|
26/6/2023 |
87 |
27 |
19 |
23 |
95 |
72 |
88 |
N |
|
27/6/2023 |
94 |
28 |
20 |
23 |
90 |
68 |
82 |
N |
|
29/6/2023 |
85 |
16 |
11 |
14 |
91 |
66 |
82 |
S – LL |
|
23/8/2023 |
90 |
32 |
20 |
25 |
94 |
55 |
76 |
N |
|
24/8/2023 |
99 |
32 |
20 |
25 |
94 |
55 |
76 |
S – LL |
|
25/8/2023 |
99 |
23 |
11 |
17 |
79 |
32 |
53 |
S – LL |
|
5/10/2023 |
<Null> |
22 |
15 |
18 |
93 |
59 |
74 |
S |
|
6/10/2023 |
85 |
24 |
14 |
18 |
87 |
45 |
70 |
S |
|
7/10/2023 |
96 |
27 |
17 |
22 |
79 |
50 |
64 |
S |
|
11/12/2023 |
89 |
29 |
18 |
24 |
91 |
53 |
75 |
N |
|
12/12/2023 |
82 |
30 |
21 |
25 |
92 |
55 |
75 |
N |
|
13/12/2023 |
<Null> |
33 |
26 |
29 |
90 |
65 |
79 |
N – LL |
Altura: Altura del dormidero, Temp Máx: temperatura máxima, Temp Mín: temperatura mínima, Temp Prom: temperatura promedio, HR Máx: humedad relativa máxima, HR Mín: humedad relativa mínima, HR Prom: humedad relativa promedio, DV y RLl: dirección del viento y registro de días con lluvia. Fuente: Los datos de temperatura y humedad relativa corresponden al data logger Hobo Pro V2 U23-002, mientras que los registros de dirección del viento y días con lluvia al Instituto Correntino del Agua y el Ambiente.
En las cartografías de los sitios dormideros por fechas y condiciones meteorológicas de las Figuras 4 N° A y B y Cuadro N° 1, en líneas generales se observan diferencias en las elecciones de los lugares dormitorio, dándose tres situaciones:
Los resultados de este estudio sugieren que la selección de árboles dormideros por Alouatta caraya está fuertemente influenciada por factores microclimáticos, lo que da respaldo a la hipótesis del confort térmico. Si bien tradicionalmente se ha planteado que primates arbóreos seleccionan árboles altos y frondosos como estrategia para evitar depredadores (Di Bitetti et al., 2000; Gazagne et al., 2025), en contextos altamente antropizados como el área de estudio, con presencia de ganadería extensiva y baja densidad de depredadores naturales, dicha hipótesis pierde fuerza. La observación de desplazamientos hacia árboles más bajos o expuestos en estas zonas sugiere que otros factores, más inmediatos y variables, como las condiciones meteorológicas, pueden incidir más decisivamente en la elección de dormideros.
Dentro del bosque, los monos encuentran un microclima más estable, con menor amplitud térmica y mayor humedad relativa. Sin embargo, este ambiente puede volverse incómodo durante días cálidos y húmedos, al limitar la disipación de calor por evapotranspiración. En este tipo de condiciones, los monos aulladores parecen preferir espacios más abiertos y elevados, posiblemente en busca de corrientes de aire que faciliten la regulación térmica. Por el contrario, en días calurosos y secos, estas áreas abiertas, como palmares y bordes de bosque, pueden ofrecer mayor ventilación, aunque aumenten su exposición a otros riesgos.
Este comportamiento podría estar vinculado también a la presencia de mosquitos y otros insectos hematófagos, que tienden a concentrarse en sitios cerrados y húmedos, como los sectores bajos del bosque. En este sentido, la elección de espacios más abiertos y ventilados en determinadas condiciones podría representar una estrategia para evitar estas molestias, que pueden afectar tanto el descanso como la salud del grupo. Cabe destacar que para la existencia de estos es necesario condiciones de altas temperaturas y humedad (Pyzczcek & Sáez Sáez, 2016), las cuales se encuentran presente en nuestra área de estudio de primavera a otoño (Contreras et al., 2020).
Cabe destacar que esta preferencia por áreas más expuestas durante eventos de calor extremo y sequía puede tener consecuencias negativas, como lo ocurrido durante los incendios forestales del 2020 (Smichowski et al., 2021), en los que se registraron altas tasas de mortalidad de monos carayá. El uso de sectores abiertos, altamente combustibles, habría aumentado su vulnerabilidad en un contexto de estrés térmico y déficit hídrico.
Finalmente, en condiciones de lluvia o viento fuerte, se observaron selecciones preferenciales de árboles más bajos, posiblemente como mecanismo de protección ante ráfagas o tormentas intensas, lo cual coincide con lo señalado en estudios previos sobre estrategias de resguardo (Gazagne et al., 2025; Brividoro et al., 2019).
En síntesis, los patrones observados en este estudio respaldan la hipótesis de que la selección de árboles dormideros por Alouatta caraya responde a una lógica de confort térmico, en función de variables meteorológicas dinámicas como temperatura, humedad, viento y precipitaciones. Este hallazgo tiene implicancias directas para la gestión del hábitat, ya que subraya la importancia de conservar áreas con diversidad estructural (en términos de altura, cobertura y volumen de copa) que permitan a los primates responder de forma flexible a condiciones ambientales cambiantes. Además, resalta la necesidad de considerar estos patrones en los planes de manejo frente a escenarios de cambio climático e incremento de eventos extremos como incendios forestales.
Conclusiones
Los resultados de este estudio aportan evidencia sustancial para considerar el confort térmico como un factor determinante en la selección de árboles dormideros por Alouatta caraya en ambientes subtropicales altamente antropizados. Si bien históricamente se ha atribuido este comportamiento a estrategias de defensa frente a depredadores o a la proximidad a recursos alimenticios, nuestras observaciones indican que las decisiones de pernocta responden también a las condiciones meteorológicas del momento, como la temperatura, la humedad, el viento o la presencia de lluvias y a la necesidad de regular térmicamente el cuerpo en un entorno cambiante. Por lo tanto, reforzamos la idea acerca de que la selección de sitios dormideros es multicausal, y que los aulladores combinan una serie de factores diarios para tomar decisiones acerca de donde dormir diferentes al azar.
La elección de sitios más abiertos y elevados en días cálidos y secos, o de sectores más protegidos y cerrados durante noches frías o lluviosas, revela una notable plasticidad conductual que permite a los monos carayá maximizar su bienestar térmico. Sin embargo, esta misma estrategia puede incrementar su vulnerabilidad frente a amenazas emergentes, como incendios forestales, especialmente cuando se ven forzados a desplazarse a zonas expuestas en busca de alivio térmico.
El uso de tecnologías de la información geográfica resulto ser clave para poder identificar y caracterizar los sitios dormideros. En este marco, el uso del drone ha sido de gran importancia, especialmente en la generación de modelos digitales de elevación de alta resolución espacial, cuales han resultado ser un gran complemento a los datos georreferenciados en campo.
Comprender estos patrones de selección espacial es clave no solo para interpretar el comportamiento de la especie, sino también para orientar políticas de conservación efectivas. La protección de hábitats con heterogeneidad estructural y climática, capaces de ofrecer múltiples opciones de resguardo frente a climas extremos, se vuelve indispensable en un contexto de cambio climático y presión antrópica creciente. Este enfoque, centrado en el bienestar térmico, puede también aplicarse a otras especies arbóreas con requerimientos ecológicos similares.
Agradecimientos
A las voluntarias Melina Ortuño, Aleksandra Nazim, Loreley Cuadrado, Yetlanezi Juans, Florencia Levit, Daniela Pucheta, Cristina Hernández, Clara Cámara, Dulce María Baumann, que acompañaron y sostuvieron el trabajo de campo durante todo el muestreo. Su compromiso, dedicación y compañía fueron fundamentales para el desarrollo de esta investigación. Sin su participación, este estudio no habría sido posible.
Referencias bibliográficas
Anderson, J. R. (1998). Sleep, sleeping sites, and sleep-related activities: Awakening to their significance. American Journal of Primatology, 46, 63–75. https://doi.org/10.1002/(SICI)1098-2345(1998)46:1<63::AID-AJP5>3.0.CO;2-T
Brividoro, M. V., Kowalewski, M. M., Scarry, C. J. y Oklander, L. I. (2019). Patterns of sleeping site and sleeping tree selection by black-and-gold howler monkeys (Alouatta caraya) in northern Argentina. International Journal of Primatology, 40(3), 374-392. 40:374–392. https://doi.org/10.1007/s10764-019-00094-x
Brividoro, M. V., Kowalewski, M. M., Scarry, C. J. y Oklander, L. I. (2019). Patterns of sleeping site and sleeping tree selection by black-and-gold howler monkeys (Alouatta caraya) in northern Argentina. Int. J. Primatol. 40, 374–392. https://doi.org/10.1007/s10764-019-00094-x
Brotcorne, F., Maslarov, C., Wandia, I. N., Fuentes, A., Beudels-Jamar, R. C. y Huynen, M. C. (2014). “The role of anthropic, ecological, and social factors in sleeping site choice by long-tailed macaques (Macaca fascicularis)”. Am. J. Primatol. 76, 1140–1150.
Contreras, F. I., Saucedo, G. I. y Smichowski, H. (2022). Análisis de la frecuencia de inundaciones en el Parque Provincial San Cayetano (Corrientes, Argentina). ScientiAmericana. Vol. 9, N°1, 35-46. http://dx.doi.org/10.30545/scientiamericana
Day, R. T., y Elwood, R. W. (1999). Sleeping site selection by the golden-handed tamarin Saguinus midas midas: the role of predation risk, proximity to feeding sites, and territorial defence. Ethology 105, 1035–1051.
Di Bitetti, M. S., Luengos Vidal, E. M., Baldovino, M. C. y Benesovsky, V. (2000). Sleeping site preferences in tufted capuchin monkeys (Cebus apella nigritus). American Journal of Primatology, 50, 257–274. https://doi.org/10.1002/(SICI)1098-2345(200004)50:4<257::AID-AJP3>3.0.CO;2-J
Di Fiore A. y Campbell C.J. (2007). The Atelines: variations in ecology, behavior, and social organization. Pp: 155 185 En: Campbell CJ, Fuentes A MacKinnon KC, Panger M. & Bearder SK, (Eds). Primates in perspective. New York: Oxford Univ. Press.
Di Fiore, A., Link, A., y Campbell, C. J. (2010). The atelines: Behavioral and socioecological diversity in a New World radiation. En: C.J. Campbell, A. Fuentes, K.C. MacKinnon, M. Panger, and S.K. Beader, (eds.) Primates in Perspective, Second Edition. Oxford: Oxford University Press.
Ellison, G., Wolfenden, A. y Kahana, L. et al. Sleeping Site Selection in the Nocturnal Northern Lesser Galago (Galago senegalensis) Supports Antipredator and Thermoregulatory Hypotheses. Int J Primatol 40, 276–296 (2019)
Eppley, T. M., Watzek, J., Dausmann, K. H., Ganzhorn, J. U. y Donati, G, (2017). Huddling is more important than rest site selection for thermoregulation in southern bamboo lemurs. Anim Behav 127:153–161. https://doi.org/10.1016/j.anbehav.2017.03.019
Fan, P. F., y Jiang, X. L. (2008). Sleeping sites, sleeping trees, and sleep-related behaviors of black crested gibbons (Nomascus concolor jingdongensis) at Mt. Wuliang, Central Yunnan, China. Am. J. Primatol. 70, 153–160.
Fei, H. L, Thompson, C. y Fan, P. F, (2019). Effects of cold weather on the sleeping behavior of Skywalker hoolock gibbons (Hoolock tianxing) in seasonal montane forest. Am J Primatol 81: e23049. https://doi.org/10.1002/ajp.23049
Gazagne, E., Goldblatt, C., Nguyễn, V. T., Brotcorne, F. y Hambuckers, A. (2025). Utilizing thermal imaging drones to investigate sleeping site selection in an arboreal primate. Current Zoology, zoae082. https://doi.org/10.1093/cz/zoae082Jardim, M. M., Queirolo, D., Peters, F. B., Mazim, F. D., Favarini, M. O., Tirelli, F. P., ... y Mourthe, I. (2020). Southern extension of the geographic range of black-and-gold howler monkeys (Alouatta caraya). Mammalia, 84(1), 102-106. doi.org/10.1515/mammalia-2018-0127
Kowalewski, M. M, Pavé R, Fernández, V. A, Raño, M. y Zunino, G. E. (2019). Life-history Traits and Group Dynamic in Black and Gold Howler Monkeys in Flooded Forests of Northern Argentina. (263-269). En: Nowak K, Barnett AA & Matsuda I, (eds.) Primates in Flooded Habitats: Ecology and Conservation. Cambridge University Press.
Kowalewski, M. M. y Gillespie T. (2018). Disturbance-tolerant primates as sentinels for global health and biodiversity conservation. (270-281). En Sanz, N.: Primatology, Biocultural Diversity and Sustainable Development in Tropical Forests, UNESCO, Mexico.
Kowalewski, M. M. (2007). Patterns of affiliation and co-operation in howler monkeys: An alternative model to explain social organization in non-human primates. Ph.D. Thesis. University of Illinois, Urbana, Illinois, USA.
Kowalewski, M., y Zunino, G. E. (2005). The parasite behavior hypothesis and the use of sleeping sites by black howler monkeys (Alouatta caraya) in a discontinuous forest. Neotropical Primates, 13(1), 22–26.
Li B-W, Li W-B, Xia D-P, Zhang T, Yang P-P y Li J-H (2022). Sleeping sites provide new insight into multiple central place foraging strategies of Tibetan macaques (Macaca thibetana). Front. Ecol. Evol. 10:1067923.
Mekonnen, A., Fashing, P.J. y Venkataraman, V.V. et al. Sleeping Site and Tree Selection by Bale Monkeys (Chlorocebus djamdjamensis) at Kokosa Forest Fragment in Southern Ethiopia. Int J Primatol 42, 915–932 (2021)
Mills, C. J, Nekaris, K. A. I., Campera, M. y Patel, E. (2021). Silky Sifakas (Propithecus candidus) Use Sleep Sites for Thermoregulation, Food Access and Predator Avoidance. Folia Primatol (Basel). 92(5-6): 315-326
Oklander, L.; Kowalewski, M.; Peker, S.; Pavé, R.; Agostini, I.; Holzmann, I. y Apellaniz, M. (2019). Alouatta carayá. En: SAyDS–SAREM (eds.) Categorización 2019 de los mamíferos de Argentina según su riesgo de extinción. Lista Roja de los mamíferos de Argentina. Versión digital: http://cma.sarem.org.ar.
Pyszczek, O. L., y Sáez Sáez, V. (2016). Ocurrencia y amenaza de Dengue, Chikungunya y Zika causada por mosquitos del género Aedes: La situación en la República Argentina 2015. Terra, 32(51), 133-161.
Rumiz DI. (1990). Alouatta caraya: Population density and demography in Northern Argentina. Am J Primatol. 21(4): 279-294.
Smichowski, H.; Montiel, M. R.; Romero, V.; Kowalewski, M. y Contreras, F.I. (2021). Evaluación de incendios en áreas periurbanas de la ciudad de corrientes (Argentina) durante la sequía extrema del año 2020. Papeles de Geografía. 67:151-167. https://doi.org/10.6018/geografia.486441
Zunino G.E. (2004). Forest disturbance effects on a population of black-and-gold howler monkeys (Alouatta caraya) in northern Argentina. ASP Bulletin 28(3): 6-7.
Zunino, G. E. (1986). Algunos aspectos de la ecología y etología del mono aullador negro (Alouatta caraya) en hábitat fragmentados (Doctoral dissertation, Universidad de Buenos Aires. Facultad de Ciencias Exactas y Naturales).
Notas
[1] Felix Ignacio Contreras, Profesor y Doctor en Geografía (Universidad Nacional del Nordeste), Investigador Adjunto de CONICET y Titular de la Cátedra Geografía Física y Geomorfología de la carrera Ingeniería en Agrimensura (UNNE). https://orcid.org/0000-0002-2548-8351
[2] Debora Rocio Gilles, Licenciada en Biología y Becaria Doctoral de CONICET y Alumna del Doctorado en Ciencias Biológicas (FaCENA-UNNE). https://orcid.org/0000-0002-0426-6215
[3] Martín Kowalewski, Licenciado en Ciencias Biológicas (UBA), Máster en Artes con orientación en Conservación Aplicada (Department of Ecology and Evolution, State University of New York at Stony Brook, USA) y Ph.D. en Antropología (University of Illinois at Urbana/Champaign (UIUC), USA). Investigador Principal de CONICET. https://orcid.org/0000-0002-6737-3771
[4] Este trabajo fue parcialmente subsidiado por los Proyectos: PI 24Q002 y 24Q004 de la Secretaría General de Ciencia y Técnica de la Universidad Nacional del Nordeste; Subsidio the American Society of Primatologists, Conservation Small Grant y Proyecto Federal de Innovación (PFI) CR 7 PFI 2023.